Ферментные препараты (пепсин, трипсин, панкреатин, лидаза, стрептолиаза и др.). Переваривание белков. Протеиназы – пепсин, трипсин, химотрипсин; проферменты протеиназ и механизмы их превращения в ферменты. Субстратная специфичность протеиназ. Экзопептидаз
Арендный блок
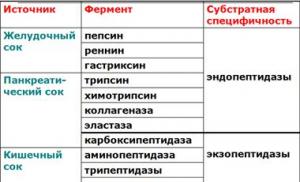
В пищевых продуктах содержание свободных аминокислот очень мало. Подавляющее их количество входит в состав белков, которые гидролизуются в ЖКТ под действием ферментов протеаз (пептидщцролаз). Субстратная специфичность этих ферментов заключается в том, что каждый из них с наибольшей скоростью расщепляет пептидные связи, образованные определёнными аминокислотами. Протеазы, гидролизующие пептидные связи внутри белковой молекулы, относят к группе эндопептидаз. Ферменты, относящиеся к группе экзопептидаз, гидролизуют пептидную связь, образованную концевыми аминокислотами. Под действием всех протеаз ЖКТ белки пищи распадаются на отдельные аминокислоты, которые затем поступают в клетки тканей.
Переваривание белков в желудке
Желудочный сок - продукт нескольких типов клеток. Обкладочные (париетальные) клетки стенок желудка образуют соляную кислоту, главные клетки секретируют пепсиноген. Добавочные и другие клетки эпителия желудка выделяют муцинсодержащую слизь. Париетальные клетки секретируют в полость желудка также гликопротеин, который называют "внутренним фактором" (фактором Касла). Этот белок связывает "внешний фактор" - витамин В12, предотвращает его разрушение и способствует всасыванию.
Образование и роль соляной кислоты. Основная пищеварительная функция желудка заключается в том, что в нём начинается переваривание белка. Существенную роль в этом процессе играет соляная кислота. Белки, поступающие в желудок, стимулируют выделение гистамина и группы белковых гормонов -гастринов которые, в свою очередь, вызывают секрецию НСI и профермента - пепсиногена. Источником Н+ является Н2СО3, которая образуется в обкладочных клетках желудка из СО2, диффундирующего из крови, и Н2О под действием фермента карбоангидразы (карбонатдегидра-тазы):
Н2О + СО2 → Н2СО3 → НСО3- + H+
Диссоциация Н2СО3 приводит к образованию бикарбоната, который с участием специальных белков выделяется в плазму в обмен на С1-, и ионов Н+, которые поступают в просвет желудка путём активного транспорта, катализируемого мембранной Н+/К+-АТФ-азой. При этом концентрация протонов в просвете желудка увеличивается в 106 раз. Ионы Сl- поступают в просвет желудка через хлоридный канал. Концентрация НСl в желудочном соке может достигать 0,16 М, за счёт чего значение рН снижается до 1,0-2,0. Приём белковой пищи часто сопровождается выделением щелочной мочи за счёт секреции большого количества бикарбоната в процессе образования НСl. Под действием НСl происходит денатурация белков пищи, не подвергшихся термической обработке, что увеличивает доступность пептидных связей для протеаз. НСl обладает бактерицидным действием и препятствует попаданию патогенных бактерий в кишечник. Кроме того, соляная кислота активирует пепсиноген и создаёт оптимум рН для действия пепсина.
Механизм активации пепсина. Под действием гастринов в главных клетках желудочных желёз стимулируются синтез и секреция пепсиногена - неактивной формы пепсина. Пепсиноген - белок, состоящий из одной полипептидной цепи с молекулярной массой 40 кД. Под действием НСl он превращается в активный пепсин (молекулярная масса 32,7 кД) с оптимумом рН 1,0-2,5. В процессе активации в результате частичного протеолиза от N-конца молекулы пепсиногена отщепляются 42 аминокислотных остатка, которые содержат почти все положительно заряженные аминокислоты, имеющиеся в пепсиногене. Таким образом, в активном пепсине преобладающими оказываются отрицательно заряженные аминокислоты, которые участвуют в конформационных перестройках молекулы и формировании активного центра. Образовавшиеся под действием НСl активные молекулы пепсина быстро активируют остальные молекулы пепсиногена (аутокатализ). Пепсин в первую очередь гидролизует пептидные связи в белках, образованные ароматическими аминокислотами (фенилаланин, триптофан, тирозин) и несколько медленнее - образованные лейцином и дикарбоновыми аминокислотами. Пепсин - эндопептидаза, поэтому в результате его действия в желудке образуются более короткие пептиды, но не свободные аминокислоты.
Переваривание белков в кишечнике.
Желудочное содержимое (химус) в процессе переваривания поступает в двенадцатиперстную кишку. Низкое значение рН химуса вызывает в кишечнике выделение белкового гормона секретина, поступающего в кровь. Этот гормон в свою очередь стимулирует выделение из поджелудочной железы в тонкий кишечник панкреатического сока, содержащего НСО3-, что приводит к нейтрализации НСl желудочного сока и ингибированию пепсина. В результате рН резко возрастает от 1,5-2,0 до ∼7,0. Поступление пептидов в тонкий кишечник вызывает секрецию другого белкового гормона - холецистокинина, который стимулирует выделение панкреатических ферментов с оптимумом рН 7,5-8,0. Под действием ферментов поджелудочной железы и клеток кишечника завершается переваривание белков.
Активация панкреатических ферментов В поджелудочной железе синтезируются проферменты ряда протеаз: трипсиноген, химотрипсиноген, проэластаза, прокарбоксипептидазы А и В. В кишечнике они путём частичного протеолиза превращаются в активные ферменты трипсин, химотрипсин, эластазу и карбоксипептидазы А и В.
Активация трипсиногена происходит под действием фермента эпителия кишечника энтеропептидазы. Этот фермент отщепляет с N-конца молекулы трипсиногена гексапептид Вал-(Асп)4-Лиз. Изменение конформации оставшейся части полипептидной цепи приводит к формированию активного центра, и образуется активный трипсин. Последовательность Вал-(Асп)4-Лиз присуща большинству известных трипсиноге-нов разных организмов - от рыб до человека.
Образовавшийся трипсин активирует химотрипсиноген, из которого получается несколько активных ферментов (рис. 9-3). Химотрипсиноген состоит из одной полипептидной цепи, содержащей 245 аминокислотных остатков и пяти дисульфидных мостиков. Под действием трипсина расщепляется пептидная связь между 15-й и 16-й аминокислотами, в результате чего образуется активный π-химотрипсин. Затем под действием π-химотрипсина отщепляется дипептид сер(14)-арг(15), что приводит к образованию δ-химотрипсина. Отщепление дипептида тре(147)-арг(148) завершает образование стабильной формы активного фермента - α-химотрипсина, который состоит из трёх полипептидных цепей, соединённых дисульфидными мостиками. Остальные проферменты панкреатических протеаз (проэластаза и прокарбоксипептидазы А и В) также активируются трипсином путём частичного протеолиза. В результате образуются активные ферменты - эластаза и карбокси-пептидазы А и В.
Специфичность действия протеаз. Трипсин преимущественно гидролизует пептидные связи, образованные карбоксильными группами аргинина и лизина. Химотрипсины наиболее активны в отношении пептидных связей, образованных карбоксильными группами ароматических аминокислот (Фен, Тир, Три). Карбоксипептидазы А и В - цинксодержащие ферменты, отщепляют С-концевые остатки аминокислот. Причём карбоксипептидаза А отщепляет преимущественно аминокислоты, содержащие ароматические или гидрофобные радикалы, а карбоксипептидаза В - остатки аргинина и лизина. Последний этап переваривания - гидролиз небольших пептидов, происходит под действием ферментов аминопептидаз и дипептидаз, которые синтезируются клетками тонкого кишечника в активной форме.
- Аминопептидазы последовательно отщепляют N-концевые аминокислоты пептидной цепи. Наиболее известна лейцинаминопептидаза - Zn2+- или Мn2+-содержащий фермент, несмотря на название, обладающий широкой специфичностью по отношению к N-концевым аминокислотам.
- Дипептидазы расщепляют дипептиды на аминокислоты, но не действуют на трипептиды.
В результате последовательного действия всех пищеварительных протеаз большинство пищевых белков расщепляется до свободных аминокислот.
Экзопептидазы (экзопротеиназы) ферменты, гидролизующие белки, отщепляя аминокислоты от конца пептида: карбоксипептидазы от C-конца, аминопептидазы от N-конца, дипептидазы расщепляют дипептиды. Экзопептидазы синтезируются в клетках тонкого кишечника (аминопептидазы, дипептидазы) и в поджелудочной железе (карбоксипептидаза). Функционируют эти ферменты внутриклеточно в кишечном эпителии и, в небольшом количестве, в просвете кишечника.
Эндопептидазы (эндопротеиназы) протеолитические ферменты (пепсин, трипсин, химотрипсин), расщепляющие пептидные связи внутри пептидной цепи. С наибольшей скоростью ими гидролизуются связи, образованные определёнными аминокислотами. Эндопептидазы синтезируются в виде проферментов, активируемых затем при помощи избирательного протеолиза. Таким образом клетки, секретирующие эти ферменты защищают собственные белки от разрушения. От действия ферментов клеточную мембрану клеток животных защищает также поверхностный слой олигосахаридов гликокаликс, а в кишечнике и желудке слой слизи.
У нас самая большая информационная база в рунете, поэтому Вы всегда можете найти походите запросы
Эта тема принадлежит разделу:
Биологическая химия
Обмен веществ в организме человека. Белки, аминокислоты, жиры. Катаболизм и анаболизм. Биохимические процессы. Предмет биологическая химия. Вопросы на экзамен и Ответы.
К данному материалу относятся разделы:
Предмет и задачи биологической химии. Обмен веществ и энергии, иерархическая структурная организация и самовоспроизведение как важнейшие признаки живой материи
Многомолекулярные системы (метаболические цепи, мембранные процессы, системы синтеза биополимеров, молекулярные регуляторные системы) как основные объекты биохимического исследования
Уровни структурной организации живого. Биохимия как молекулярный уровень изучения явлений жизни. Биохимия и медицина (медицинская биохимия)
Основные разделы и направления в биохимии: биоорганическая химия, динамическая и функциональная биохимия, молекулярная биология
История изучения белков. Представление о белках как важнейшем классе органических веществ и структурно-функциональном компоненте организма человека
Аминокислоты, входящие в состав белков, их строение и свойства. Пептидная связь. Первичная структура белков
Зависимость биологических свойств белков от первичной структуры. Видовая специфичность первичной структуры белков (инсулины разных животных)
Конформация пептидных цепей в белках (вторичная и третичная структуры). Слабые внутримолекулярные взаимодействия в пептидной цепи; дисульфидные связи
Основы функционирования белков. Активный центр белков и его специфическое взаимодействие с лигандом как основа биологической функции всех белков. Комплементарность взаимодействия молекул белка с лигандом. Обратимость связывания
Доменная структура и её роль в функционировании белков. Яды и лекарства как ингибиторы белков
Четвертичная структура белков. Особенности строения и функционирования олигомерных белков на примере гемсодержащего белка - гемоглобина
Лабильность пространственной структуры белков и их денатурация. Факторы, вызывающие денатурацию
Шапероны - класс белков, защищающий другие белки от денатурации в условиях клетки и облегчающий формирование их нативной конформации
Многообразие белков. Глобулярные и фибриллярные белки, простые и сложные. Классификация белков по их биологическим функциям и по семействам: (сериновые протеазы, иммуноглобулины)
Иммуноглобулины, особенности строения, избирательность взаимодействия с антигеном. Многообразие антигенсвязывающих участков Н- и L-цепей. Классы иммуноглобулинов, особенности строения и функционирования
Физико-химические свойства белков. Молекулярный вес, размеры и форма, растворимость, ионизация, гидратация
Методы выделения индивидуальных белков: осаждение солями и органическими растворителями, гель-фильтрация, электрофорез, ионообменная и аффинная хроматография
Методы количественного измерения белков. Индивидуальные особенности белкового состава органов. Изменения белкового состава органов при онтогенезе и болезнях.
История открытия и изучения ферментов. Особенности ферментативного катализа. Специфичность действия ферментов. Зависимость скорости ферментативных реакций от температуры, рН, концентрации фермента и субстрата.
Классификация и номенклатура ферментов. Изоферменты. Единицы измерения активности и количества ферментов.
Кофакторы ферментов: ионы металлов и коферменты. Коферментные функции витаминов (на примере витаминов В6, РР, В2)
Ингибиторы ферментов. Обратимое и необратимое ингибирование. Конкурентное ингибирование. Лекарственные препараты как ингибиторы ферментов.
Регуляция действия ферментов: аллостерические ингибиторы и активаторы. Каталитический и регуляторный центры. Четвертичная структура аллостерических ферментов и кооперативные изменения конформации протомеров фермента.
Регуляция активности ферментов путем фосфорилирования и дефосфорилирования. Участие ферментов в проведении гормонального сигнала
Различия ферментного состава органов и тканей. Органоспецифические ферменты. Изменение ферментов в процессе развития
Изменение активности ферментов при болезнях. Наследственные энзимопатии. Происхождение ферментов крови и значение их определения при болезнях
Применение ферментов для лечения болезней. Применение ферментов как аналитических реагентов при лабораторной диагностике (определении глюкозы, этанола, мочевой кислоты и т.д.). Иммобилизованные ферменты
Обмен веществ: питание, метаболизм и выделение продуктов метаболизма. Органические и минеральные компоненты пищи. Основные и минорные компоненты
Основные пищевые вещества: углеводы, жиры, белки, суточная потребность, переваривание; частичная взаимозаменяемость при питании
Незаменимые компоненты основных пищевых веществ. Незаменимые аминокислоты; пищевая ценность различных пищевых белков. Линолевая кислота - незаменимая жирная кислота
История открытия и изучения витаминов. Классификация витаминов. Функции витаминов.
Алиментарные и вторичные авитаминозы и гиповитаминозы. Гипервитаминозы. Примеры
Минеральные вещества пищи. Региональные патологии, связанные с недостаточностью микроэлементов в пище и воде.
Понятие о метаболизме и метаболических путях. Ферменты и метаболизм. Понятие о регуляции метаболизма. Основные конечные продукты метаболизма у человека
Исследования на целых организмах, органах, срезах тканей, гомогенатах, субклеточных структурах и на молекулярном уровне
Эндэргонические и экзэргонические реакции в живой клетке. Макроэргические соединения. Примеры.
Окислительное фосфорилирование, коэффициент Р/О. Строение митохондрий и структурная организация дыхательной цепи. Трансмембранный электрохимический потенциал.
Регуляция цепи переноса электронов (дыхательный контроль). Разобщение тканевого дыхания и окислительного фосфорилирования. Терморегуляторная функция тканевого дыхания
Нарушения энергетического обмена: гипоэнергетические состояния как результат гипоксии, гипо-, авитаминозов и других причин. Возрастная характеристика энергетического обеспечения организма питательными веществами
Образование токсических форм кислорода, механизм их повреждающего действия на клетки. Механизмы устранения токсичных форм кислорода
Катаболизм основных пищевых веществ - углеводов, жиров, белков. Понятие о специфических путях катаболизма и общих путях катаболизма
Окислительное декарбоксилирование пировиноградной кислоты. Последовательность реакций. Строение пируватдекарбоксилазного комплекса
Цикл лимонной кислоты: последовательность реакций и характеристика ферментов. Связь между общими путями катаболизма и цепью переноса электронов и протонов
Механизмы регуляции цитратного цикла. Анаболические функции цикла лимонной кислоты. Реакции, пополняющие цитратный цикл
Основные углеводы животных, их содержание в тканях, биологическая роль. Основные углеводы пищи. Переваривание углеводов
Аэробный распад основной путь катаболизма глюкозы у человека и других аэробных организмов. Последовательность реакций до образования пирувата (аэробный гликолиз)
Распространение и физиологическое значение аэробного распада глюкозы. Использование глюкозы для синтеза жиров в печени и в жировой ткани.
Анаэробный распад глюкозы (анаэробный гликолиз). Гликолитическая оксиредукция, пируват как акцептор водорода. Субстратное фосфорилирование. Распространение и физиологическое значение этого пути распада глюкозы
Биосинтез глюкозы (глюконеогенез) из аминокислот, глицерина и молочной кислоты. Взаимосвязь гликолиза в мышцах и глюконеогенеза в печени (цикл Кори)
Представление о пентозофосфатном пути превращений глюкозы. Окислительные реакции (до стадии рибулозо-5-фосфата). Распространение и суммарные результаты этого пути (образование пентоз, НАДФН и энергетика)
Свойства и распространение гликогена как резервного полисахарида. Биосинтез гликогена. Мобилизация гликогена
Особенности обмена глюкозы в разных органах и клетках: эритроциты, мозг, мышцы, жировая ткань, печень.
Представление о строении и функциях углеводной части гликолипидов и гликопротеинов. Сиаловые кислоты
Наследственные нарушения обмена моносахаридов и дисахаридов: галактоземия, непереносимость фруктозы и дисахаридов. Гликогенозы и агликогенозы
Важнейшие липиды тканей человека. Резервные липиды (жиры) и липиды мембран (сложные липиды). Жирные кислоты липидов тканей человека.
Незаменимые факторы питания липидной природы. Эссенциальные жирные кислоты: ω-3- и ω-6-кислоты как предшественники синтеза эйкозаноидов.
Биосинтез жирных кислот, регуляция метаболизма жирных кислот
Химизм реакций β-окисления жирных кислот, энергетический итог
Пищевые жиры и их переваривание. Всасывание продуктов переваривания. Нарушение переваривания и всасывания. Ресинтез триацилглицеринов в стенке кишечника
Образование хиломикронов и транспорт жиров. Роль апопротеинов в составе хиломикронов. Липопротеинлипаза
Биосинтез жиров в печени из углеводов. Структура и состав транспортных липопротеинов крови
Депонирование и мобилизация жиров в жировой ткани. Регуляция синтеза и мобилизации жиров. Роль инсулина, глюкагона и адреналина
Основные фосфолипиды и гликолипиды тканей человека (глицерофосфолипиды, сфингофосфолипиды, гликоглицеролипиды, гликосфиголипиды). Представление о биосинтезе и катаболизме этих соединений.
Нарушение обмена нейтрального жира (ожирение), фосфолипидов и гликолипидов. Сфинголипидозы
Строение и биологические функции эйкозаноидов. Биосинтез простагландинов и лейкотриенов
Холестерин как предшественник ряда других стероидов. Представление о биосинтезе холестерина. Написать ход реакций до образования мевалоновой кислоты. Роль гидроксиметилглутарил-КоА-редуктазы
Синтез желчных кислот из холестерина. Конъюгация желчных кислот, первичные и вторичные желчные кислоты. Выведение желчных кислот и холестерина из организма.
ЛПНП и ЛПВП - транспортные, формы холестерина в крови, роль в обмене холестерина. Гиперхолестеринемия. Биохимические основы развития атеросклероза.
Механизм возникновения желчнокаменной болезни (холестериновые камни). Применение хенодезокеихолевой кислоты для лечения желчнокаменной болезни.
Переваривание белков. Протеиназы - пепсин, трипсин, химотрипсин; проферменты протеиназ и механизмы их превращения в ферменты. Субстратная специфичность протеиназ. Экзопептидазы и эндопептидазы.
Диагностическое значение биохимического анализа желудочного и дуоденального сока. Дать краткую характеристику состава этих соков.
Протеиназы поджелудочной железы и панкреатиты. Применение ингибиторов протеиназ для лечения панкреатитов.
Трансаминирование: аминотрансферазы; коферментная функция витамина В6. Специфичность аминотрансфераз
Аминокислоты, участвующие в трансаминировании; особая роль глутаминовой кислоты. Биологическое значение реакций трансаминирования. Определение трансаминаз в сыворотке крови при инфаркте миокарда и болезнях печени.
Окислительное дезаминирование аминокислот; глутаматдегидрогеназа. Непрямое дезаминирование аминокислот. Биологическое значение.
Глутаминаза почек; образование и выведение солей аммония. Активация глутаминазы почек при ацидозе
Биосинтез мочевины. Связь орнитинового цикла с ЦТК. Происхождение атомов азота мочевины. Нарушения синтеза и выведения мочевины. Гипераммонемии
Обмен безазотистого остатка аминокислот. Гликогенные и кетогенные аминокислоты. Синтез глюкозы из аминокислот. Синтез аминокислот из глюкозы
Трансметилирование. Метионин и S-аденозилметионин. Синтез креатина, адреналина и фосфатидилхолинов
Метилирование ДНК. Представление о метилировании чужеродных и лекарственных соединений
Антивитамины фолиевой кислоты. Механизм действия сульфаниламидных препаратов.
Обмен фенилаланина и тирозина. Фенилкетонурия; биохимический дефект, проявление болезни, методы предупреждения, диагностика и лечение.
Алкаптонурия и альбинизм: биохимические дефекты, при которых они развиваются. Нарушение синтеза дофамина, паркинсонизм
Декарбоксилирование аминокислот. Структура биогенных аминов (гистамин, серотонин, γ-аминомасляная кислота, катехоламины). Функции биогенных аминов
Дезаминирование и гидроксилирование биогеных аминов (как реакции обезвреживания этих соединений)
Нуклеиновые кислоты, химический состав, строение. Первичная структура ДНК и РНК, связи, формирующие первичную структуру
Вторичная и третичная структура ДНК. Денатурация, ренативация ДНК. Гибридизация, видовые различия первичной структуры ДНК
РНК, химический состав, уровни структурной организации. Типы РНК, функции. Строение рибосомы.
Строение хроматина и хромосомы
Распад нуклеиновых кислот. Нуклеазы пищеварительного тракта и тканей. Распад пуриновых нуклеотидов.
Представление о биосинтезе пуриновых нуклеотидов; начальные стадии биосинтеза (от рибозо-5-фосфата до 5-фосфорибозиламина)
Инозиновая кислота как предшественник адениловой и гуаниловой кислот.
Представление о распаде и биосинтезе пиримидиновых нуклеотидов
Нарушения обмена нуклеотидов. Подагра; применение аллопуринола для лечения подагры. Ксантинурия. Оротацидурия
Биосинтез дезоксирибонуклеотидов. Применение ингибиторов синтеза дезоксирибонуклеотидов для лечения злокачественных опухолей
Синтез ДНК и фазы клеточного деления. Роль циклинов и циклинзависимых протеиназ в продвижении клетки по клеточному циклу
Повреждение и репарация ДНК. Ферменты ДНК-репарирующего комплекса
Биосинтез РНК. РНК полимеразы. Понятие о мозаичной структуре генов, первичном транскрипте, посттранскрипционном процессинге
Биологический код, понятия, свойства кода, коллинеарность, сигналы терминации.
Роль транспортных РНК в биосинтезе белков. Биосинтез аминоацил-т-РНК. Субстратная специфичность аминоацил-т-РНК-синтетаз.
Последовательность событий на рибосоме при сборке полипептидной цепи. Функционирование полирибосом. Посттрансляционный процессинг белков
Адаптивная регуляция генов у про- и эукариотов. Теория оперона. Функционирование оперонов
Понятие о клеточной дифференцировке. Изменение белкового состава клеток при дифференцировке (на примере белкового состава полипептидных цепей гемоглобина)
Молекяулрные механизмы генетической изменчивости. Молекулярные мутации: типы, частота, значение
Генетическая гетерогенность. Полиморфизм белков в популяции человека (варианты гемоглобина, гликозилтрансферазы, группоспецифических веществ и др)
Биохимические основы возникновения и проявления наследственных болезней (разнообразие, распространение)
Основные системы межклеточной коммуникации: эндокринная, паракринная, аутокринная регуляция
Роль гормонов в системе регуляции метаболизма. Клетки-мишени и клеточные рецепторы гормонов
Механизмы передачи гормональных сигналов в клетки
Классификация гормонов по химическому строению и биологическим функциям
Строение, синтез и метаболизм иодтиронинов. Влияние на обмен веществ. Изменение метаболизма при гипо- и гипертиреозе. Причины и проявление эндемического зоба
Регуляция энергетического метаболизма, роль инсулина и контринсулярных гормонов в обеспечении гомеостаза
Изменения метаболизма при сахарном диабете. Патогенез основных симптомов сахарного диабета
Патогенез поздних осложнений сахарного диабета (макро- и микроангиопатии, нефропатия, ретинопатия, катаракта). Диабетическая кома
Регуляция водно-солевого обмена. Строение и функции альдостерона и вазопрессина
Система ренин-ангиотензин-альдостерон. Биохимические механизмы возникновения почечной гипертонии, отеков, дегидратации.
Токсичность кислорода: образование активных форм кислорода (супероксид анион, перекись водорода, гидроксильный радикал)
Повреждение мембран в результате перекисного окисления липидов. Механизмы защиты от токсического действия кислорода: неферментативные (витамины Е, С, глутатион и др.) и ферментативные (супероксиддисмутаза, каталаза, глутатионпероксидаза)
Биотрансформация лекарственных веществ. Влияние лекарств на ферменты, участвующие в обезвреживании ксенобиотиков
Основы химического канцерогенеза. Представление о некоторых химических канцерогенах: полициклические ароматические углеводороды, ароматические амины, диоксиды, митоксины, нитрозамины
Особенности развития, строения и метаболизма эритроцитов
Транспорт кислорода и диоксида углерода кровью. Гемоглобин плода (HbF) и его физиологическое значение
Полиморфные формы гемоглобинов человека. Гемоглобинопатии. Анемические гипоксии
Биосинтез гема и его регуляция. Нарушения синтеза тема. Порфирии
Распад гема. Обезвреживание билирубина. Нарушения обмена билирубинажелтухи: гемолитическая, обтурационная, печеночно-клеточная. Желтуха новорожденных
Диагностическое значение определения билирубина и других желчных пигментов в крови и моче
Обмен железа: всасывание, транспорт кровью, депонирование. Нарушение обмена железа: железодефицитная анемия, гемохроматоз
Основные белковые фракции плазмы крови и их функции. Значение их определения для диагностики заболеваний. Энзимодиагностика
Свертывающая система крови. Этапы образования фибринового сгустка. Внутренний и внешний пути свертывания и их компоненты
Принципы образования и последовательность фукционирования ферментных комплексов прокоагулянтного пути. Роль витамина К в свертывании крови
Основные механизмы фибринолиза. Активаторы плазминогена как тромболитические средства. Основаные антикоагулянты крови: антитромбин III, макроглобулин, антиконвертин. Гемофилии.
Клиническое значение биохимического анализа крови
Основные мембраны клетки и их функции. Общие свойства мембран: жидкостность, поперечная асимметрия, избирательная проницаемость
Липидный состав мембран (фосфолипиды, гликолипиды, холестерин). Роль липидов в формировании липидного бислоя
Белки мембран - интегральные, поверхностные, «заякоренные». Значение посттрансляционных модификаций в образовании функциональных мембранных белков
Механизмы переноса веществ через мембраны: простая диффузия, первично-активный транспорт (Nа+-К+-АТФаза, Са2+-АТФаза), пассивный симпорт и антипорт, вторично-активный транспорт
Трансмембранная передача сигнала. Участие мембран в активации внутриклеточных регуляторных систем - аденилатциклазной и инозитолфосфатной в передаче гормонального сигнала
Коллаген: особенности аминокислотного состава, первичной и пространственной структуры. Роль аскорбиновой кислоты в гидоксилировании пролина и лизина
Особенности биосинтеза и созревания коллагена. Проявления недостаточности витамина С
Особенности строения и функции эластина
Гликозаминогликаны и протеогликаны. Строение и функции. Роль гиалуроновой кислоты в организации межклеточного матрикса
Адгезивные белки межклеточного матрикса: фибронектин и ламинин, их строение и функции. Роль этих белков в межклеточных взаимодействиях и развитии опухолей
Структурная организация межклеточного матрикса. Изменения соединительной ткани при старении, коллагенозах. Роль коллагеназы при заживлении ран. Оксипролинурия
Важнейшие белки миофибрилл: миозин, актин, актомиозин, тропомиозин, тропонин, актинин. Молекулярная структура миофибрилл
Биохимические механизмы мышечного сокращения и расслабления. Роль градиента одновалентных ионов и ионов кальция в регуляции мышечного сокращения и расслабления
Саркоплазматические белки: миоглобин, его строение и функции. Экстрактивные вещества мышц
Особенности энергетического обмена в мышцах. Креатинфосфат
Биохимические изменения при мышечных дистрофиях и денервации мышц. Креатинурия
Химический состав нервной ткани. Миелиновые мембраны: особенности состава и структуры
Энергетический обмен в нервной ткани. Значение аэробного распада глюкозы
Биохимия возникновения и проведения нервного импульса. Молекулярные механизмы синаптической передачи
Медиаторы: ацетилхолин, катехоламины, серотонин, γ-аминомаслянная кислота, глутаминовая кислота, глицин, гистамин
Энерго- и ресурсосбережение
Гелиоколекторы Расчет гелиоколлектора Определение площади солнечных коллекторов. Главным достоинством солнечных коллекторов является то, что вырабатываемая ими тепловая энергия бесплатна.
История XVII-XIX века
Физическая культура, спорт и оздоровление
Физическое воспитание в оздоровительном лагере имеет специфические особенности, обусловленные сравнительно коротким периодом пребыванием в лагере, разнообразием контингента детей по возрасту, состоянию здоровья, уровню физического развития и физической подготовленности.
Маркетинг промислового підприємства
Сутність і функції маркетингу промислового підприємства. Товари промислової продукції ТПП. Маркетингові дослідження на ринку товарів промислової продукції ТПП. Ціноутворення на нову продукцію.
Воєнна стратегія Богдана Хмельницького
Визвольна війна українського народу середини 17 ст. сформувала в Україні цілу когорту талановитих воєначальників. Богдан Хмельницький мав воєнно-стратегічну мету, що включала такі основні напрямки. Битва під Пилявцями. Воєнна кампанія.
Переваривание происходит под действием протеаз - пептидгидролаз. Протеазы, гидролизирующие пептидные связи внутри молекулы - эндопептидазы, концевые аминокислоты - экзопептидазы.
Специфичность действия протеаз. Трипсин преимущественно гидролизует пептидные связи, образованные карбоксильными группами аргинина и лизина. Химотрипсины наиболее активны в отношении пептидных связей, образованных карбоксильными группами ароматических аминокислот. Карбоксипептидазы А и В - цинксодержащие ферменты, отщепляют С-концевые остатки аминокислот. Причём карбоксипептидаза А отщепляет преимущественно аминокислоты, содержащие ароматические или гидрофобные радикалы, а карбоксипептидаза В - остатки аргинина и лизина. Последний этап переваривания - гидролиз небольших пептидов, происходит под действием ферментов аминопептидаз и дипептидаз, которые синтезируются клетками тонкого кишечника в активной форме.
Дипептидазы расщепляют дипептиды на аминокислоты, но не действуют на трипептиды.
В результате последовательного действия всех пищеварительных протеаз большинство пищевых белков расщепляется до свободных аминокислот.
Эндопептидазы (эндопротеиназы) протеолитические ферменты (пепсин, трипсин, химотрипсин), расщепляющие пептидные связи внутри пептидной цепи. С наибольшей скоростью ими гидролизуются связи, образованные определёнными аминокислотами.
Экзопептидазы (экзопротеиназы) ферменты, гидролизующие белки, отщепляя аминокислоты от конца пептида: карбоксипептидазы от C-конца, аминопептидазы от N-конца, дипептидазы расщепляют дипептиды. Экзопептидазы синтезируются в клетках тонкого кишечника (аминопептидазы, дипептидазы) и в поджелудочной железе (карбоксипептидаза). Функционируют эти ферменты внутриклеточно в кишечном эпителии и, в небольшом количестве, в просвете кишечника.
Экзопептидазы отщепляют концевые амикислоты, освобождая их от бремени пептидной связи, VIVA LA RESISTANCE!!!
Пепсиноген - белок, состоящий из одной полипептидной цепи с молекулярной массой 40 кД. Под действием НСl он превращается в активный пепсин (с оптимумом рН 1,0-2,5. В процессе активации в результате частичного протеолиза от N-конца молекулы пепсиногена отщепляются 42 аминокислотных остатка, которые содержат почти все положительно заряженные аминокислоты, имеющиеся в пепсиногене. Таким образом, в активном пепсине преобладающими оказываются отрицательно заряженные аминокислоты, которые участвуют в конформационных перестройках молекулы и формировании активного центра.
Активация панкреатических ферментов. В поджелудочной железе синтезируются проферменты ряда протеаз: трипсиноген, химотрипсиноген, проэластаза, прокарбоксипептидазы А и В. В кишечнике они путём частичного протеолиза превращаются в активные ферменты трипсин, химотрипсин, эластазу и карбоксипептидазы А и В.
Активация трипсиногена происходит под действием фермента эпителия кишечника энтеропептидазы. Этот фермент отщепляет с N-конца молекулы трипсиногена гексапептид Вал-(Асп)4-Лиз. Изменение конформации оставшейся части полипептидной цепи приводит к формированию активного центра, и образуется активный трипсин. Последовательность Вал-(Асп)4-Лиз присуща большинству известных трипсиноге-нов разных организмов - от рыб до человека.
(?) 76. Диагностическое значение биохимического анализа желудочного и дуоденального сока. Дать краткую характеристику состава этих соков.
Желудочный сок сложный по составу пищеварительный сок, вырабатываемый различными клетками слизистой оболочки желудка. Желудочный сок содержит соляную кислоту и ряд минеральных солей, а также различные ферменты, главнейшими из которых являются пепсин, расщепляющий белки, химозин (сычужный фермент), створаживающий молоко, липаза, расщепляющая жиры. Составной частью желудочного сока является также слизь, играющая важную роль в защите слизистой оболочки желудка от раздражающих веществ, попавших в него; при высокой кислотности желудочного сока слизь нейтрализует ее.Кроме соляной кислоты, ферментов, солей и слизи, в желудочном соке содержится также особое вещество т. наз. внутренний фактор Касла. Это вещество необходимо для всасывания витамина В12 в тонких кишках, что обеспечивает нормальное созревание красных кровяных телец в костном мозге. При отсутствии фактора Касла в желудочном соке, что обычно связано с заболеванием желудка, а иногда с его оперативным удалением, развивается тяжелая форма малокровия. Анализ желудочного сока является очень важным методом исследования больных с заболеваниями желудка, кишечника, печени, желчного пузыря, крови и пр
Дуоденальный сок - пищеварительный сок двенадцатиперстной кишки, состоящий из секрета поджелудочной железы, желчи, сока кишечных крипт и дуоденальных желез.
(?) 77. Протеиназы поджелудочной железы и панкреатиты. Применение ингибиторов протеиназ для лечения панкреатитов.

Панкреатит - это воспаление поджелудочной железы. Болезнь может протекать в острой (быстро и бурно) или хронической (долго и вяло) форме, с периодами обострения хронического панкреатита.
Причины панкреатита
Употребление алкоголя и заболевания желчного пузыря (прежде всего, желчнокаменная болезнь) в 95-98% случаев являются причинами панкреатита.
Другие факторы риска, которые могут спровоцировать воспаление поджелудочной железы:
В норме в поджелудочной железе вырабатываются неактивные предшественники ферментов - их переход в активную форму происходит непосредственно в 12-перстной кишке, куда они поступают по протоку поджелудочной железы и общему желчному протоку.
Под действием различных факторов (например, камень закупоривающий желчный проток) повышается давление в протоке поджелудочной железы, нарушается отток ее секрета, и происходит преждевременная активация ферментов. В результате вместо того, чтобы переваривать пищу, ферменты начинают переваривать саму поджелудочную железу. Развивается острое воспаление.
При хроническом панкреатите нормальная ткань поджелудочной железы постепенно замещается рубцовой, развивается недостаточность экзокринной (выработка ферментов) и эндокринной (выработка гормонов, в том числе, инсулина) функций железы.
В 1930 г. Фрей открыл первый ингибитор калликреина. В последующем это вещество было получено в чистом виде и применено с лечебной целью. В клинической практике для лечения острого панкреатита широкое распространение получили ингибиторы протеаз трасилол, контрикал, тцалол, пантрипин и др. Трасилол является полипептидом с молекулярной массой 11 600, состоящим из 18 аминокислот. Он игибирует калликреин, трипсин, химотрипсин и плазмин путем образования с ферментами неактивного комплекса. Кроме того, трасилол и другие ингибиторы протеаз дают выраженный вазопрессорный эффект и, таким образом, имеют значение в предупреждении шока. Под влиянием трасилола, по мнению разных авторов, быстро купируется болевой синдром, снижаются токсемия и симптомы шока. При назначении больших доз одного из ингибиторов протеаз мы также в большинстве случаев наблюдали улучшение состояния тяжелобольных (исчезновение болей и др.). Однако лечение всегда было комплексным и трудно сказать, насколько в этих случаях помогали именно ингибиторы протеаз.
- 2. Гетеротрофные и аутотрофные организмы: различия по питанию и источникам энергии. Катаболизм и анаболизм.
- 3. Многомолекулярные системы (метаболические цепи, мембранные процессы, системы синтеза биополимеров, молекулярные регуляторные системы) как основные объекты биохимического исследования.
- 4. Уровни структурной организации живого. Биохимия как молекулярный уровень изучения явлений жизни. Биохимия и медицина (медицинская биохимия).
- 5. Основные разделы и направления в биохимии: биоорганическая химия, динамическая и функциональная биохимия, молекулярная биология.
- 6. История изучения белков. Представление о белках как важнейшем классе органических веществ и структурно-функциональном компоненте организма человека.
- 7. Аминокислоты, входящие в состав белков, их строение и свойства. Пептидная связь. Первичная структура белков.
- 8. Зависимость биологических свойств белков от первичной структуры. Видовая специфичность первичной структуры белков (инсулины разных животных).
- 9. Конформация пептидных цепей в белках (вторичная и третичная структуры). Слабые внутримолекулярные взаимодействия в пептидной цепи; дисульфидные связи.
- 11. Доменная структура и её роль в функционировании белков. Яды и лекарства как ингибиторы белков.
- 12.Четвертичная структура белков. Особенности строения и функционирования олигомерных белков на примере гемсодержащего белка - гемоглобина.
- 13.Лабильность пространственной структуры белков и их денатурация. Факторы, вызывающие денатурацию.
- 14.Шапероны - класс белков, защищающий другие белки от денатурации в условиях клетки и облегчающий формирование их нативной конформации.
- 15.Многообразие белков. Глобулярные и фибриллярные белки, простые и сложные. Классификация белков по их биологическим функциям и по семействам: (сериновые протеазы, иммуноглобулины).
- 17.Физико-химические свойства белков. Молекулярный вес, размеры и форма, растворимость, ионизация, гидратация
- 18.Методы выделения индивидуальных белков: осаждение солями и органическими растворителями, гель-фильтрация, электрофорез, ионообменная и аффинная хроматография.
- 19.Методы количественного измерения белков. Индивидуальные особенности белкового состава органов. Изменения белкового состава органов при онтогенезе и болезнях.
- 21 .Классификация и номенклатура ферментов. Изоферменты. Единицы измерения активности и количества ферментов.
- 22.Кофакторы ферментов: ионы металлов и коферменты. Коферментные функции витаминов (на примере витаминов в6, рр, в2).
- 23.Ингибиторы ферментов. Обратимое и необратимое ингибирование. Конкурентное ингибирование. Лекарственные препараты как ингибиторы ферментов.
- 25.Регуляция активности ферментов путем фосфорилирования и дефосфорилирования. Участие ферментов в проведении гормонального сигнала.
- 26.Различия ферментного состава органов и тканей. Органоспецифические ферменты. Изменение ферментов в процессе развития.
- 27.Изменение активности ферментов при болезнях. Наследственные энзимопатии. Происхождение ферментов крови и значение их определения при болезнях.
- 29.Обмен веществ: питание, метаболизм и выделение продуктов метаболизма. Органические и минеральные компоненты пищи. Основные и минорные компоненты.
- 30.Основные пищевые вещества: углеводы, жиры, белки, суточная потребность, переваривание; частичная взаимозаменяемость при питании.
- 31 .Незаменимые компоненты основных пищевых веществ. Незаменимые аминокислоты; пищевая ценность различных пищевых белков. Линолевая кислота - незаменимая жирная кислота.
- 32.История открытия и изучения витаминов. Классификация витаминов. Функции витаминов.
- 34.Минеральные вещества пищи. Региональные патологии, связанные с недостаточностью микроэлементов в пище и воде.
- 35.Понятие о метаболизме и метаболических путях. Ферменты и метаболизм. Понятие о регуляции метаболизма. Основные конечные продукты метаболизма у человека
- 36.Исследования на целых организмах, органах, срезах тканей, гомогенатах, субклеточных структурах и на молекулярном уровне
- 37.Эндэргонические и экзэргонические реакции в живой клетке. Макроэргические соединения. Примеры.
- 39.Окислительное фосфорилирование, коэффициент р/о. Строение митохондрий и структурная организация дыхательной цепи. Трансмембранный электрохимический потенциал.
- 40.Регуляция цепи переноса электронов (дыхательный контроль). Разобщение тканевого дыхания и окислительного фосфорилирования. Терморегуляторная функция тканевого дыхания
- 42.Образование токсических форм кислорода, механизм их повреждающего действия на клетки. Механизмы устранения токсичных форм кислорода.
- 43.Катаболизм основных пищевых веществ - углеводов, жиров, белков. Понятие о специфических путях катаболизма и общих путях катаболизма.
- 44.Окислительное декарбоксилирование пировиноградной кислоты. Последовательность реакций. Строение пируватдекарбоксилазного комплекса.
- 45.Цикл лимонной кислоты: последовательность реакций и характеристика ферментов. Связь между общими путями катаболизма и цепью переноса электронов и протонов.
- 46.Механизмы регуляции цитратного цикла. Анаболические функции цикла лимонной кислоты. Реакции, пополняющие цитратный цикл
- 47.Основные углеводы животных, их содержание в тканях, биологическая роль. Основные углеводы пищи. Переваривание углеводов
- 49. Аэробный распад - основной путь катаболизма глюкозы у человека и других аэробных организмов. Последовательность реакций до образования пирувата (аэробный гликолиз).
- 50.Распространение и физиологическое значение аэробного распада глюкозы. Использование глюкозы для синтеза жиров в печени и в жировой ткани.
- 52. Биосинтез глюкозы (глюконеогенез) из аминокислот, глицерина и молочной кислоты. Взаимосвязь гликолиза в мышцах и глюконеогенеза в печени (цикл Кори).
- 54. Свойства и распространение гликогена как резервного полисахарида. Биосинтез гликогена. Мобилизация гликогена.
- 55. Особенности обмена глюкозы в разных органах и клетках: эритроциты, мозг, мышцы, жировая ткань, печень.
- 56. Представление о строении и функциях углеводной части гликолипидов и гликопротеинов. Сиаловые кислоты
- 57. Наследственные нарушения обмена моносахаридов и дисахаридов: галактоземия, непереносимость фруктозы и дисахаридов. Гликогенозы и агликогенозы
- Глицеральдегид -3 –фосфат
- 58. Важнейшие липиды тканей человека. Резервные липиды (жиры) и липиды мембран (сложные липиды). Жирные кислоты липидов тканей человека.
- Состав жирных кислот подкожного жира человека
- 59. Незаменимые факторы питания липидной природы. Эссенциальные жирные кислоты: ω-3- и ω-6-кислоты как предшественники синтеза эйкозаноидов.
- 60.Биосинтез жирных кислот, регуляция метаболизма жирных кислот
- 61.Химизм реакций β-окисления жирных кислот, энергетический итог.
- 6З.Пищевые жиры и их переваривание. Всасывание продуктов переваривания. Нарушение переваривания и всасывания. Ресинтез триацилглицеринов в стенке кишечника.
- 64.Образование хиломикронов и транспорт жиров. Роль апопротеинов в составе хиломикронов. Липопротеинлипаза.
- 65.Биосинтез жиров в печени из углеводов. Структура и состав транспортных липопротеинов крови.
- 66. Депонирование и мобилизация жиров в жировой ткани. Регуляция синтеза и мобилизации жиров. Роль инсулина, глюкагона и адреналина.
- 67.Основные фосфолипиды и гликолипиды тканей человека (глицерофосфолипиды, сфингофосфолипиды, гликоглицеролипиды, гликосфиголипиды). Представление о биосинтезе и катаболизме этих соединений.
- 68.Нарушение обмена нейтрального жира (ожирение), фосфолипидов и гликолипидов. Сфинголипидозы
- Сфинголипиды, метаболизм: заболевания сфинголипидозы, таблица
- 69.Строение и биологические функции эйкозаноидов. Биосинтез простагландинов и лейкотриенов.
- 70.Холестерин как предшественник ряда других стероидов. Представление о биосинтезе холестерина. Написать ход реакций до образования мевалоновой кислоты. Роль гидроксиметилглутарил-КоА-редуктазы.
- 71.Синтез желчных кислот из холестерина. Конъюгация желчных кислот, первичные и вторичные желчные кислоты. Выведение желчных кислот и холестерина из организма.
- 72.Лпнп и лпвп - транспортные, формы холестерина в крови, роль в обмене холестерина. Гиперхолестеринемия. Биохимические основы развития атеросклероза.
- 73. Механизм возникновения желчнокаменной болезни (холестериновые камни). Применение хенодезокеихолевой кислоты для лечения желчнокаменной болезни.
- 75. Переваривание белков. Протеиназы - пепсин, трипсин, химотрипсин; проферменты протеиназ и механизмы их превращения в ферменты. Субстратная специфичность протеиназ. Экзопептидазы и эндопептидазы.
- 76. Диагностическое значение биохимического анализа желудочного и дуоденального сока. Дать краткую характеристику состава этих соков.
- 77. Протеиназы поджелудочной железы и панкреатиты. Применение ингибиторов протеиназ для лечения панкреатитов.
- 78. Трансаминирование: аминотрансферазы; коферментная функция витамина в6. Специфичность аминотрансфераз.
- 80. Окислительное дезаминирование аминокислот; глутаматдегидрогеназа. Непрямое дезаминирование аминокислот. Биологическое значение.
- 82. Глутаминаза почек; образование и выведение солей аммония. Активация глутаминазы почек при ацидозе.
- 83. Биосинтез мочевины. Связь орнитинового цикла с цтк. Происхождение атомов азота мочевины. Нарушения синтеза и выведения мочевины. Гипераммонемии.
- 84. Обмен безазотистого остатка аминокислот. Гликогенные и кетогенные аминокислоты. Синтез глюкозы из аминокислот. Синтез аминокислот из глюкозы.
- 85. Трансметилирование. Метионин и s-аденозилметионин. Синтез креатина, адреналина и фосфатидилхолинов
- 86. Метилирование днк. Представление о метилировании чужеродных и лекарственных соединений.
- 88. Антивитамины фолиевой кислоты. Механизм действия сульфаниламидных препаратов.
- 89. Обмен фенилаланина и тирозина. Фенилкетонурия; биохимический дефект, проявление болезни, методы предупреждения, диагностика и лечение.
- 90. Алкаптонурия и альбинизм: биохимические дефекты, при которых они развиваются. Нарушение синтеза дофамина, паркинсонизм.
- 91. Декарбоксилирование аминокислот. Структура биогенных аминов (гистамин, серотонин, γ-аминомасляная кислота, катехоламины). Функции биогенных аминов.
- 92. Дезаминирование и гидроксилирование биогеных аминов (как реакции обезвреживания этих соединений).
- 93. Нуклеиновые кислоты, химический состав, строение. Первичная структура днк и рнк, связи, формирующие первичную структуру
- 94. Вторичная и третичная структура днк. Денатурация, ренативация днк. Гибридизация, видовые различия первичной структуры днк.
- 95. Рнк, химический состав, уровни структурной организации. Типы рнк, функции. Строение рибосомы.
- 96. Строение хроматина и хромосомы
- 97. Распад нуклеиновых кислот. Нуклеазы пищеварительного тракта и тканей. Распад пуриновых нуклеотидов.
- 98. Представление о биосинтезе пуриновых нуклеотидов; начальные стадии биосинтеза (от рибозо-5-фосфата до 5-фосфорибозиламина).
- 99. Инозиновая кислота как предшественник адениловой и гуаниловой кислот.
- 100. Представление о распаде и биосинтезе пиримидиновых нуклеотидов.
- 101. Нарушения обмена нуклеотидов. Подагра; применение аллопуринола для лечения подагры. Ксантинурия. Оротацидурия.
- 102. Биосинтез дезоксирибонуклеотидов. Применение ингибиторов синтеза дезоксирибонуклеотидов для лечения злокачественных опухолей.
- 104. Синтез днк и фазы клеточного деления. Роль циклинов и циклинзависимых протеиназ в продвижении клетки по клеточному циклу.
- 105. Повреждение и репарация днк. Ферменты днк-репарирующего комплекса.
- 106. Биосинтез рнк. Рнк полимеразы. Понятие о мозаичной структуре генов, первичном транскрипте, посттранскрипционном процессинге.
- 107. Биологический код, понятия, свойства кода, коллинеарность, сигналы терминации.
- 108. Роль транспортных рнк в биосинтезе белков. Биосинтез аминоацил-т-рнк. Субстратная специфичность аминоацил-т-рнк-синтетаз.
- 109. Последовательность событий на рибосоме при сборке полипептидной цепи. Функционирование полирибосом. Посттрансляционный процессинг белков.
- 110. Адаптивная регуляция генов у про- и эукариотов. Теория оперона. Функционирование оперонов.
- 111. Понятие о клеточной дифференцировке. Изменение белкового состава клеток при дифференцировке (на примере белкового состава полипептидных цепей гемоглобина).
- 112. Молекяулрные механизмы генетической изменчивости. Молекулярные мутации: типы, частота, значение
- 113. Генетическая гетерогенность. Полиморфизм белков в популяции человека (варианты гемоглобина, гликозилтрансферазы, группоспецифических веществ и др).
- 114. Биохимические основы возникновения и проявления наследственных болезней (разнообразие, распространение).
- 115. Основные системы межклеточной коммуникации: эндокринная, паракринная, аутокринная регуляция.
- 116. Роль гормонов в системе регуляции метаболизма. Клетки-мишени и клеточные рецепторы гормонов
- 117. Механизмы передачи гормональных сигналов в клетки.
- 118. Классификация гормонов по химическому строению и биологическим функциям
- 119. Строение, синтез и метаболизм иодтиронинов. Влияние на обмен веществ. Изменение метаболизма при гипо- и гипертиреозе. Причины и проявление эндемического зоба.
- 120. Регуляция энергетического метаболизма, роль инсулина и контринсулярных гормонов в обеспечении гомеостаза.
- 121. Изменения метаболизма при сахарном диабете. Патогенез основных симптомов сахарного диабета.
- 122. Патогенез поздних осложнений сахарного диабета (макро- и микроангиопатии, нефропатия, ретинопатия, катаракта). Диабетическая кома.
- 123. Регуляция водно-солевого обмена. Строение и функции альдостерона и вазопрессина
- 124. Система ренин-ангиотензин-альдостерон. Биохимические механизмы возникновения почечной гипертонии, отеков, дегидратации.
- 125. Роль гормонов в регуляции обмена кальция и фосфатов (паратгормон, кальцитонин). Причины и проявления гипо- и гиперпаратироидизма.
- 126. Строение, биосинтез и механизм действия кальцитриола. Причины и проявление рахита
- 127. Строение и секреция кортикостероидов. Изменения катаболизма при гипо- и гиперкортицизме.
- 128. Регуляция синтезами секреции гормонов по принципу обратной связи.
- 129. Половые гормоны: строение, влияние на обмен веществ и функции половых желез, матки и молочных желез.
- 130. Гормон роста, строение, функции.
- 131. Метаболизм эндогенных и чужеродных токсических веществ: реакции микросомального окисления и реакции конъюгации с глутатионом, глюкуроновой кислотой, серной кислотой.
- 132. Металлотионеин и обезвреживание ионов тяжелых металлов. Белки теплового шока.
- 133. Токсичность кислорода: образование активных форм кислорода (супероксид анион, перекись водорода, гидроксильный радикал).
- 135. Биотрансформация лекарственных веществ. Влияние лекарств на ферменты, участвующие в обезвреживании ксенобиотиков.
- 136. Основы химического канцерогенеза. Представление о некоторых химических канцерогенах: полициклические ароматические углеводороды, ароматические амины, диоксиды, митоксины, нитрозамины.
- 137. Особенности развития, строения и метаболизма эритроцитов.
- 138. Транспорт кислорода и диоксида углерода кровью. Гемоглобин плода (HbF) и его физиологическое значение.
- 139. Полиморфные формы гемоглобинов человека. Гемоглобинопатии. Анемические гипоксии
- 140. Биосинтез гема и его регуляция. Нарушения синтеза тема. Порфирии.
- 141. Распад гема. Обезвреживание билирубина. Нарушения обмена билирубина-желтухи: гемолитическая, обтурационная, печеночно-клеточная. Желтуха новорожденных.
- 142. Диагностическое значение определения билирубина и других желчных пигментов в крови и моче.
- 143. Обмен железа: всасывание, транспорт кровью, депонирование. Нарушение обмена железа: железодефицитная анемия, гемохроматоз.
- 144. Основные белковые фракции плазмы крови и их функции. Значение их определения для диагностики заболеваний. Энзимодиагностика.
- 145. Свертывающая система крови. Этапы образования фибринового сгустка. Внутренний и внешний пути свертывания и их компоненты.
- 146. Принципы образования и последовательность фукционирования ферментных комплексов прокоагулянтного пути. Роль витамина к в свертывании крови.
- 147. Основные механизмы фибринолиза. Активаторы плазминогена как тромболитические средства. Основаные антикоагулянты крови: антитромбин III, макроглобулин, антиконвертин. Гемофилии.
- 148. Клиническое значение биохимического анализа крови.
- 149. Основные мембраны клетки и их функции. Общие свойства мембран: жидкостность, поперечная асимметрия, избирательная проницаемость.
- 150. Липидный состав мембран (фосфолипиды, гликолипиды, холестерин). Роль липидов в формировании липидного бислоя.
- 151. Белки мембран - интегральные, поверхностные, «заякоренные». Значение посттрансляционных модификаций в образовании функциональных мембранных белков.
- 153. Трансмембранная передача сигнала. Участие мембран в активации внутриклеточных регуляторных систем - аденилатциклазной и инозитолфосфатной в передаче гормонального сигнала.
- 154. Коллаген: особенности аминокислотного состава, первичной и пространственной структуры. Роль аскорбиновой кислоты в гидоксилировании пролина и лизина.
Переваривание белков в желудке
Желудочный сок - продукт нескольких типов клеток. Обкладочные (париетальные) клетки стенок желудка образуют соляную кислоту, главные клетки секретируют пепсиноген. Добавочные и другие клетки эпителия желудка выделяют муцинсодержащую слизь. Париетальные клетки секретируют в полость желудка также гликопротеин, который называют "внутренним фактором" (фактором Касла). Этот белок связывает "внешний фактор" - витамин В 12 , предотвращает его разрушение и способствует всасыванию.
Образование и роль соляной кислоты . Основная пищеварительная функция желудка заключается в том, что в нём начинается переваривание белка. Существенную роль в этом процессе играет соляная кислота. Белки, поступающие в желудок, стимулируют выделение гистамина и группы белковых гормонов -гастринов которые, в свою очередь, вызывают секрецию НСI и профермента - пепсиногена. Источником Н + является Н 2 СО 3 , которая образуется в обкладочных клетках желудка из СО 2 , диффундирующего из крови, и Н 2 О под действием фермента карбоангидразы (карбонатдегидра-тазы):
Н 2 О + СО 2 → Н 2 СО 3 → НСО 3 - + H +
Диссоциация Н 2 СО 3 приводит к образованию бикарбоната, который с участием специальных белков выделяется в плазму в обмен на С1 - , и ионов Н + , которые поступают в просвет желудка путём активного транспорта, катализируемого мембранной Н + /К + -АТФ-азой. При этом концентрация протонов в просвете желудка увеличивается в 10 6 раз. Ионы Сl - поступают в просвет желудка через хлоридный канал. Концентрация НСl в желудочном соке может достигать 0,16 М, за счёт чего значение рН снижается до 1,0-2,0. Приём белковой пищи часто сопровождается выделением щелочной мочи за счёт секреции большого количества бикарбоната в процессе образования НСl. Под действием НСl происходит денатурация белков пищи, не подвергшихся термической обработке, что увеличивает доступность пептидных связей для протеаз. НСl обладает бактерицидным действием и препятствует попаданию патогенных бактерий в кишечник. Кроме того, соляная кислота активирует пепсиноген и создаёт оптимум рН для действия пепсина.
Механизм активации пепсина . Под действием гастринов в главных клетках желудочных желёз стимулируются синтез и секреция пепсиногена - неактивной формы пепсина. Пепсиноген - белок, состоящий из одной полипептидной цепи с молекулярной массой 40 кД. Под действием НСl он превращается в активный пепсин (молекулярная масса 32,7 кД) с оптимумом рН 1,0-2,5. В процессе активации в результате частичного протеолиза от N-конца молекулы пепсиногена отщепляются 42 аминокислотных остатка, которые содержат почти все положительно заряженные аминокислоты, имеющиеся в пепсиногене. Таким образом, в активном пепсине преобладающими оказываются отрицательно заряженные аминокислоты, которые участвуют в конформационных перестройках молекулы и формировании активного центра. Образовавшиеся под действием НСl активные молекулы пепсина быстро активируют остальные молекулы пепсиногена (аутокатализ). Пепсин в первую очередь гидролизует пептидные связи в белках, образованные ароматическими аминокислотами (фенилаланин, триптофан, тирозин) и несколько медленнее - образованные лейцином и дикарбоновыми аминокислотами. Пепсин - эндопептидаза, поэтому в результате его действия в желудке образуются более короткие пептиды, но не свободные аминокислоты.
Переваривание белков в кишечнике .
Желудочное содержимое (химус) в процессе переваривания поступает в двенадцатиперстную кишку. Низкое значение рН химуса вызывает в кишечнике выделение белкового гормона секретина, поступающего в кровь. Этот гормон в свою очередь стимулирует выделение из поджелудочной железы в тонкий кишечник панкреатического сока, содержащего НСО 3 - , что приводит к нейтрализации НСl желудочного сока и ингибированию пепсина. В результате рН резко возрастает от 1,5-2,0 до ∼7,0. Поступление пептидов в тонкий кишечник вызывает секрецию другого белкового гормона - холецистокинина, который стимулирует выделение панкреатических ферментов с оптимумом рН 7,5-8,0. Под действием ферментов поджелудочной железы и клеток кишечника завершается переваривание белков.
Активация панкреатических ферментов В поджелудочной железе синтезируются проферменты ряда протеаз: трипсиноген, химотрипсиноген, проэластаза, прокарбоксипептидазы А и В. В кишечнике они путём частичного протеолиза превращаются в активные ферменты трипсин, химотрипсин, эластазу и карбоксипептидазы А и В.
Активация трипсиногена происходит под действием фермента эпителия кишечника энтеропептидазы. Этот фермент отщепляет с N-конца молекулы трипсиногена гексапептид Вал-(Асп) 4 -Лиз. Изменение конформации оставшейся части полипептидной цепи приводит к формированию активного центра, и образуется активный трипсин. Последовательность Вал-(Асп) 4 -Лиз присуща большинству известных трипсиноге-нов разных организмов - от рыб до человека.
Образовавшийся трипсин активирует химотрипсиноген , из которого получается несколько активных ферментов (рис. 9-3). Химотрипсиноген состоит из одной полипептидной цепи, содержащей 245 аминокислотных остатков и пяти дисульфидных мостиков. Под действием трипсина расщепляется пептидная связь между 15-й и 16-й аминокислотами, в результате чего образуется активный π-химотрипсин. Затем под действием π-химотрипсина отщепляется дипептид сер(14)-арг(15), что приводит к образованию δ-химотрипсина. Отщепление дипептида тре(147)-арг(148) завершает образование стабильной формы активного фермента - α-химотрипсина, который состоит из трёх полипептидных цепей, соединённых дисульфидными мостиками. Остальные проферменты панкреатических протеаз (проэластаза и прокарбоксипептидазы А и В) также активируются трипсином путём частичного протеолиза. В результате образуются активные ферменты - эластаза и карбокси-пептидазы А и В.
Специфичность действия протеаз . Трипсин преимущественно гидролизует пептидные связи, образованные карбоксильными группами аргинина и лизина. Химотрипсины наиболее активны в отношении пептидных связей, образованных карбоксильными группами ароматических аминокислот (Фен, Тир, Три). Карбоксипептидазы А и В - цинксодержащие ферменты, отщепляют С-концевые остатки аминокислот. Причём карбоксипептидаза А отщепляет преимущественно аминокислоты, содержащие ароматические или гидрофобные радикалы, а карбоксипептидаза В - остатки аргинина и лизина. Последний этап переваривания - гидролиз небольших пептидов, происходит под действием ферментов аминопептидаз и дипептидаз, которые синтезируются клетками тонкого кишечника в активной форме.
Аминопептидазы последовательно отщепляют N-концевые аминокислоты пептидной цепи. Наиболее известна лейцинаминопептидаза - Zn 2+ - или Мn 2+ -содержащий фермент, несмотря на название, обладающий широкой специфичностью по отношению к N-концевым аминокислотам.
Дипептидазы расщепляют дипептиды на аминокислоты, но не действуют на трипептиды.
В результате последовательного действия всех пищеварительных протеаз большинство пищевых белков расщепляется до свободных аминокислот.
Экзопептидазы (экзопротеиназы) -ферменты, гидролизующие белки, отщепляяаминокислотыот концапептида:карбоксипептидазы- от C-конца,аминопептидазы- от N-конца,дипептидазырасщепляют дипептиды. Экзопептидазы синтезируются в клеткахтонкого кишечника(аминопептидазы, дипептидазы) и вподжелудочной железе(карбоксипептидаза). Функционируют эти ферменты внутриклеточно в кишечномэпителиии, в небольшом количестве, в просветекишечника.
Эндопептидазы (эндопротеиназы) -протеолитические ферменты(пепсин,трипсин,химотрипсин), расщепляющиепептидные связивнутрипептидной цепи. С наибольшей скоростью ими гидролизуются связи, образованные определённымиаминокислотами. Эндопептидазы синтезируются в видепроферментов, активируемых затем при помощи избирательногопротеолиза. Таким образом клетки, секретирующие эти ферменты защищают собственные белки от разрушения. От действия ферментовклеточную мембрануклеток животных защищает также поверхностный слойолигосахаридов-гликокаликс, а вкишечникеижелудке- слой слизи.
ПЕПСИН - основной протеолитический фермент желудочного сока (КФ 3.4.23.1), относится к группе пептид-гидролаз, расщепляет белки, в основном до полипептидов, хотя среди продуктов расщепления белков пепсином встречаются низкомолекулярные пептиды и аминокислоты. Препараты П. используют в качестве лекарственных средств для заместительной терапии при расстройствах пищеварения. При нек-рых патол, состояниях активность П. в желудочном соке (см.) является одним из диагностических признаков и определяется в клинико-биологических лабораториях. Содержание профермента П. - пепсиногена в моче (уропепсина) - служит дополнительным диагностическим тестом при исследовании секреторной способности слизистой оболочки желудка. П. находит применение в пищевой и мясо-молочной промышленности.
П. открыт в 1836 г. Т. Шванном, а в 1930 г. получен Нортропом (J. Н. Northrop) в кристаллическом виде.
П. является наиболее хорошо изученным представителем подкласса карбоксильных протеиназ (см. Пептид-гидролазы). Мол. вес (масса) П. ок. 35 000, изоэлектрическая точка (см.) находится при pH ниже 1,0, что обусловлено высоким содержанием в молекуле фермента остатков аспарагиновой и глутаминовой к-т при низком содержании диаминокислот, а также присутствием одного остатка фосфорной к-ты. Молекула П. состоит из одиночной полипептидной цепи из 327 аминокислотных остатков и представляет собой глобулу с размерами осей 5,5 X 4,5 X 3,2 нм, составленную из двух доменов, имеющих сходную структуру. Для молекулы П. характерно очень низкое содержание альфа-спиральных участков и высокое содержание бета-структур. Между доменами находится впадина, в к-рой располагается активный центр П., он формируется аминокислотными остатками, локализованными в разных доменах; каталитическими группами являются COOH-группы остатков аспарагиновой к-ты в 32-м и 215-м положениях.
Активность П., как и других карбоксильных протеиназ, подавляется диазокарбонильными ингибиторами и нек-рыми эпоксидами, специфически блокирующими COOH-группы активного центра фермента. Природным ингибитором П. является пепстатин - N-замещенный пентапептид, продуцируемый нек-рыми стрептомицетами.
П. наиболее устойчив при pH ок. 5,0-5,5. В более кислой среде происходит самопереваривание (аутолиз) фермента; при pH выше 6,0 наступает его быстрая и практически необратимая инактивация. П. инактивируется также при температуре выше 60°.
П. содержится в желудочном соке млекопитающих, птиц, рептилий и рыб. У беспозвоночных и микроорганизмов обнаружены ферменты, близкие по свойствам к П. В желудочном соке человека и высших млекопитающих наряду с П. присутствует гастриксин - фермент, имеющий сходные с П. свойства и гомологичную структуру.
Синтезируется П. главными клетками желез слизистой оболочки желудка (см.) в виде неактивного предшественника - профермента пепсиногена, к-рый в присутствии соляной к-ты желудочного сока превращается в активный фермент. При этом в результате конформационных изменений и гидролиза пептидной связи между лей44-иле45 от N-концевого участка молекулы пепсиногена отщепляется фрагмент, и т. о. образуется активный П., катализирующий затем аутокаталитическую активацию следующих порций профермента. Один из отщепляющихся при этом пептидов, так наз. ингибитор П. с мол. весом (массой) ок. 3000, при pH выше 5,0 подавляет активность П.; при pH ниже 4,0 ингибитор быстро расщепляется. В моче млекопитающих, в т. ч. и человека, в норме обнаруживается пепсиноген (уропепсин), проникающий в мочу из главных клеток слизистой оболочки желудка через кровь и почки (см. Уропепсин).
Процесс переваривания белков в жел.-киш. тракте начинается с действия П. Этот фермент обладает широкой субстратной специфичностью; он катализирует в белках гидролиз пептидных связей, образованных различными аминокислотными остатками. П. расщепляет почти все белки растительного и животного происхождения, за исключением протаминов и кератинов. Оптимум действия П. при pH 2,0. При pH ок. 5,0 П. створаживает молоко, вызывая превращение казеиногена в казеин (см.). П. способен гидролизовать ряд низкомолекулярных синтетических пептидов и эфиров, в состав к-рых входят ароматические аминокислоты. Оптимум для гидролиза П. многих синтетических субстратов находится при pH 4,0. П. катализирует также реакцию транс-пептидирования (перенос аминокислотного остатка с одного синтетического субстрата на другой).
Коммерческие препараты неочищенного П. получают из кислотного экстракта (аутолизата) слизистой оболочки желудка путем высаливания 15% р-ром NaCl и последующего высушивания. Очищенный П. выделяют из таких препаратов с помощью ионообменной хроматографии (см.). Аналог П. - химозин (см.), используемый в пищевой промышленности для сыроварения, получают таким же способом из слизистой оболочки сычуга - отдела желудка крупного рогатого скота. Для мед. целей применяют П. из слизистой оболочки желудка свиней. Кристаллический П. может быть получен как из коммерческих препаратов П., так и из желудочного сока или непосредственно из слизистой оболочки желудка.
Для определения активности П. предложено много методов. Ранее использовали метод Метта, к-рый, однако, устарел и не дает точных результатов. Наиболее часто для определения активности П. применяют метод Ансона - расщепление под действием П. денатурированного гемоглобина с последующим определением количества тирозина в без-белковом фильтрате (см. Ансона - Черникова метод). Для исследования активности П. в желудочном соке и содержания уропепсина в моче широко применяется метод Пятницкого, основанный на определении створаживающей активности фермента.
При ряде заболеваний жел.-киш. тракта - хрон, гастрите (см.), язве желудка и двенадцатиперстной кишки (см. Язвенная болезнь), раке желудка (см.) и при нек-рых патол, состояниях - пернициозной анемии (см.), гипохромной анемии (см. Железодефицитная анемия) нарушается секреция П. В связи с этим определение П. в желудочном соке наряду с определением концентрации соляной к-ты имеет диагностическое значение. Для диагностических целей используют также определение в моче уропепсина, содержание к-рого, как полагают, отражает уровень секреторной способности слизистой оболочки желудка.
Пепсин как лекарственный препарат
Препарат П. (Pepsinum), используемый как лекарственное средство, получают из слизистой оболочки желудка свиней, в качестве наполнителя применяют сахарозу или лактозу. Препарат представляет собой белый или кремовый порошок сладкого вкуса со специфическим запахом, растворимый в воде, в 20% этиловом спирте и нерастворимый в эфире и хлороформе.
Обычно препараты П. обладают довольно низкой протеолитической активностью: 1 г препарата содержит всего 5 мг чистого фермента.
Для обеспечения оптимального действия препарата реакция среды в желудке должна быть кислой, а концентрация свободной соляной к-ты - не ниже 0,15-0,2%.
П. применяют для заместительной терапии при расстройствах пищеварения (см.), сопровождающихся секреторной недостаточностью желудка (ахилии, гипацидных и анацидных гастритах, диспепсиях и др.). Следует учитывать, что эндогенный П., как и другие пищеварительные ферменты, главные клетки слизистой обрлочки желудка выделяют обычно в избыточных количествах, поэтому снижение переваривающей способности желудочного сока при снижении его кислотности часто является результатом недостаточного выделения соляной к-ты, а не снижения активности или интенсивности образования П. Т. о., при гипацидных состояниях основное значение имеет обеспечение оптимальных условий для переваривания желудочного содержимого, а применение П. имеет вспомогательное значение. При анацидных состояниях, когда снижена кислотообразующая функция желудка, целесообразно назначать П. в сочетании с разведенной соляной к-той.
П. назначают внутрь: взрослым по 0,2-0,5 г на прием 2-3 раза в день перед едой или во время еды в порошках или в 1-3% р-ре соляной к-ты (10-15 капель на полстакана воды); детям на прием назначают по 0,05-0,3 г в 0,5-1% р-ре соляной к-ты. Противопоказаниями к приему П. являются гиперацидный гастрит, обострение при язве желудка. Побочным действием препарат, применяемый в терапевтических дозах, не обладает.
Форма выпуска: порошок. Хранение: в хорошо укупоренных банках в прохладном (от 2 до 15°), защищенном от света месте.
Кроме препарата пепсина (Pepsinum), фармацевтическая промышленность выпускает препарат ацидин-пепсин (Acidin-pepsinum), содержащий 1 часть пепсина и 4 части бетаина гидрохлорида (см. Бетаины), к-рый в желудке гидролизуется с образованием свободной соляной к-ты (0,4 г бетаина соответствуют примерно 16 каплям разведенной соляной к-ты). Таблетки ацидин-пепсина (по 0,5 и 0,25 г) растворяют в половине стакана воды и принимают 3-4 раза в день во время еды.
За рубежом таблетки, содержащие П., выпускаются под названиями «Ацидол-пепсин», «Бетацид», «Аци-пепсол», «Пепсамин».
Библиография: Андреева Н. С. и др. Рентгеноструктурный анализ пепсина, Молек. биол., т. 12, № 4, с. 922, 1978, библиогр.; Лобарева Л. С. и Степанов В. М. Карбоксильные протеиназы плесневых грибов, в кн.: Усп. биол, хим., под ред. Б. Н. Степаненко, т. 19, с. 83, М., 1978, библиогр.; Мосолов В. В. Протеолитические ферменты, с. 101 и др., М., 1971; Нортроп Д., Кунитц М. и Херриотт Р. Кристаллические ферменты, пер. с англ., с. 32, М., 1950; Радбиль О. С. Фармакологические основы лечения болезней органов пищеварения, с. 78, М., 1976; Acid proteases, structure, function and biology, ed. by J. Tang, N. Y., 1977; Tang J. Evolution in the structure and function of carboxyl proteases, Molec, cell. Biochem., v. 26, p. 93, 1979.
Л. А. Локшина; H. В. Коробов (фарм.).
В виде неактивного предшественника (профермента) трипсиногена . Трипсины ряда животных получены в кристаллическом виде (впервые в 1932). Молекула бычьего трипсина (молекулярная масса около 24 кДа) состоит из 223 аминокислотных остатков, образующих одну полипептидную цепь, и содержит 6 дисульфидных связей. Изоэлектрическая точка трипсина лежит при 10,8, а оптимум каталитической активности - при pH 7,8-8,0.
Трипсины относятся к группе сериновых протеаз и содержат в активном центре остатки серина и гистидина . Трипсины легко подвергаются самоперевариванию (аутолизу), что приводит к загрязнению препаратов трипсинов неактивными продуктами (промышленный препарат содержит до 50 % неактивных примесей). Препараты трипсина высокой чистоты получают хроматографическими методами.
Физические свойства
Трипсин представляет собой бесцветное кристаллическое вещество с температурой плавления около 150.
Биологические свойства и функции
Основной функцией является пищеварение. Катализирует гидролиз белков и пептидов. Может находиться в неактивном состоянии в виде трипсиногена. Активируется, в том числе кишечным ферментом энтеропептидазой , путем отщепления гексапептида. Катализирует также гидролиз восков-сложных эфиров. Оптимум каталитической активности - при 7,8-8. Активный центр имеет белковую природу и состоит в основном из серина и гистидина . Синтезируется в виде трипсиногена в поджелудочной железе и используется в кишечнике млекопитающих и рыб . Превращает остальные проферменты гидролаз в активные ферменты .
Применение
Трипсин используют для изготовления лекарств . Препараты трипсина обладают противовоспалительным и противоотёчным действием (при внутривенном и внутримышечном введении); способны избирательно расщеплять ткани, подвергшиеся некрозу . В медицине трипсин применяют для лечения ран, ожогов, тромбозов, часто в сочетании с другими ферментами и с антибиотиками . Трипсин входит в состав препаратов для системной энзимотерапии - вобэнзима , флогэнзима.
Литература
- Нортроп Д., Кунитц М., Херриотт Р. Кристаллические ферменты, пер. с англ., М., 1950;
- Мосолов В. В. . Протеолитические ферменты, М., 1971.
- В. И. Стручков, А. В. Григорян, В. К. Гостищев, С. В. Лохвицкий, Л. С. Тапинский Протеолитические ферменты в гнойной хирургии., М., 1970 г.;
- Проблемы медицинской энзимологии» (под редакцией С. Р. Мардашова) . М., 1970 г.;
- К. Н. Веремеенко Протеолитические ферменты поджелудочной железы и их применение в клинике. Киев, 1967 г.
- Системная энзимотерапия. Опыт и перспективы / Под ред. В.И. Кулакова, В.А. Насоновой, В.С. Савельева. – СПб.: Интер-Медика. 2004. – 264 с.
- Тец В.В., Кнорринг Г.Ю., Артеменко Н.К. и др. Влияние экзогенных протеолитических ферментов на бактерии // Антибиотики и химиотерапия. – 2004. – Т. 49. – № 12. – С. 9–13.
- Полиферментные препараты в гнойной хирургии: Методические рекомендации / Под ред. член–корр. РАМН Н.А. Ефименко. – М., 2005. – 32 с.
Родственные ферменты
«Ближайшие родственники» трипсина - трипсиноген , пепсин , химотрипсин. У высших животных обнаружены анионные аналоги трипсина, у многих животных и растений-нейтральные.
Изменение гидролитических свойств
Активность трипсина подавляется фосфорорганическими соединениями, некоторыми металлами, а также рядом высокомолекулярных белковых веществ - ингибиторов трипсина, содержащихся в тканях животных, растений и микроорганизмов. Ионы 2+ , 2+ , 2+ , 2+ , 2+ повышают гидролитическую активность трипсина.
Ссылки
| Эндопептидазы : сериновые протеазы/сериновые эндопептидазы (КФ 3.4.21) | |
|---|---|
| Ферменты пищеварения | Энтеропептидаза - Трипсин - Химотрипсин - Эластаза (Нейтрофильная, Панкреатическая) |
| Коагуляция | факторы:
Тромбин - Фактор VIIa - Фактор IXa - Фактор Xa - Фактор XIa - Фактор XIIa - Калликреин (PSA) фибринолиз : Плазмин - Активатор плазминогена (Тканевой - Урокиназный) |
| Система комплемента | Фактор B - Фактор D - Фактор I - MASP (MASP1, MASP2) - C3-конвертаза |
| Другие, иммунная система : | Химаза - Гранзим - Триптаза - Протеиназа 3/Миелобластин |
| Из биотоксинов: | Анкрод - Батроксобин |
| Остальные | Акрозин - Пролилэндопептидаза - Проназа - Пропротеинконвертаза (1, 2) - Рилин - Субтилизин/Фурин - Стрептокиназа - S1P - Катепсин ( , ) |
Wikimedia Foundation . 2010 .
Синонимы :Смотреть что такое "Трипсин" в других словарях:
- (Тrypsinum). Эндогенный протеолитический фермент, разрываюший пептидные связи в молекуле белка. Расщепляет также высокомолекулярные продукты распада белков, полипептиды типа пептонов, а также некоторые низкомолекулярные пептиды, содержащие… … Словарь медицинских препаратов
ТРИПСИН - ТРИПСИН, фермент, входящий в состав поджелудочного сока, описан Кюне (Kiihne) в 1867 г., относится к группе протеаз. Он расщепляет белки,протамины,пептоны и полипептиды. В наиболее чистом виде, свободном от липазы и амилазы, получен по способу… … Большая медицинская энциклопедия
ТРИПСИН - Trypsinum. Свойства. Получают из поджелудочной железы крупного рогатого скота. Это белый или белый с желтоватым оттенком порошок или пористая масса без запаха. Легко растворим в воде и изотоническом растворе хлорида натрия; рН 0,2 % водного раст … Отечественные ветеринарные препараты
Фермент поджелудочной железы, растворяющий белок. Словарь иностранных слов, вошедших в состав русского языка. Чудинов А.Н., 1910. трипсин (гр. thrypsis разжижение) пищеварительный фермент, синтезируемый в поджелудочной железе в виде неактивного… … Словарь иностранных слов русского языка
ТРИПСИН, пищеварительный фермент, выделяемый ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗОЙ. Выделяется в неактивной форме (чтобы не повредить ткани на пути в кишечник), затем фермент тонкой кишки преобразует его в активный трипсин. Расщепляет пептидные связи аминокислот … Научно-технический энциклопедический словарь
Пищеварительный фермент, вырабатываемый в виде неактивного трипсиногена поджелудочной железой человека и животных; расщепляет белки в кишечнике … Большой Энциклопедический словарь
ТРИПСИН, трипсина, мн. нет, муж. (от греч. tripsis размалывание) (биол.). Вещество сока поджелудочной железы, переводящее белки в более простые, растворимые соединения. Толковый словарь Ушакова. Д.Н. Ушаков. 1935 1940 … Толковый словарь Ушакова
Протеолитич. фермент, синтезируемый клетками поджелудочной железы в форме неактивного предшественника трипсиногена. Активирует проферменты поджелудочной железы и занимает ключевое положение в пищеварении в тонком отделе кишечника. В кристаллич.… … Биологический энциклопедический словарь
Сущ., кол во синонимов: 4 лекарство (1413) протеаза (4) фермент (253) … Словарь синонимов
трипсин
- Один из основных пищеварительных (протеолитических) ферментов, катализирующий гидролиз пептидов и др. соединений по месту связей, в которых участвуют карбоксильные группы аргинина и лизина }